Blas M. Benito
Senior Data Scientist & Team Lead
AgTech
Bio
I’m a senior data scientist & team lead with 19+ years of international experience bridging cutting-edge research and production systems. I build geospatial and machine learning pipelines into scalable solutions to solve hard problems and drive business impact.
Current Role
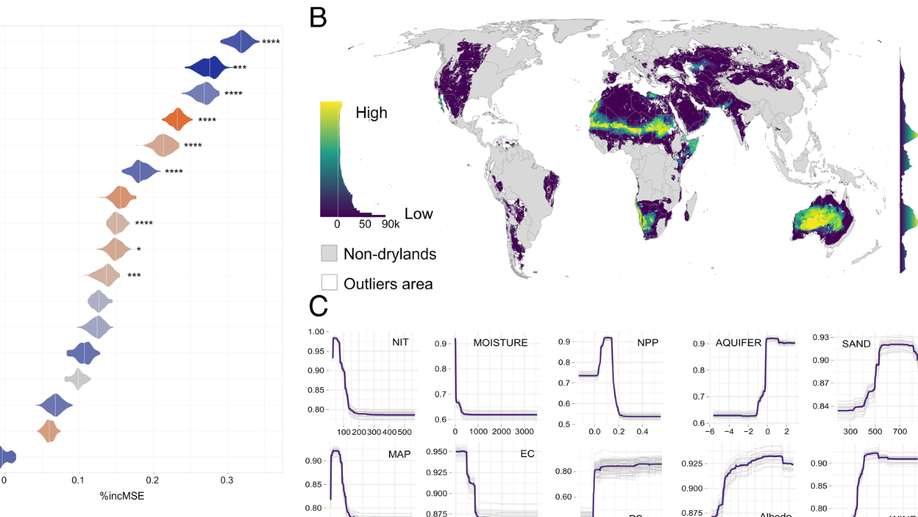
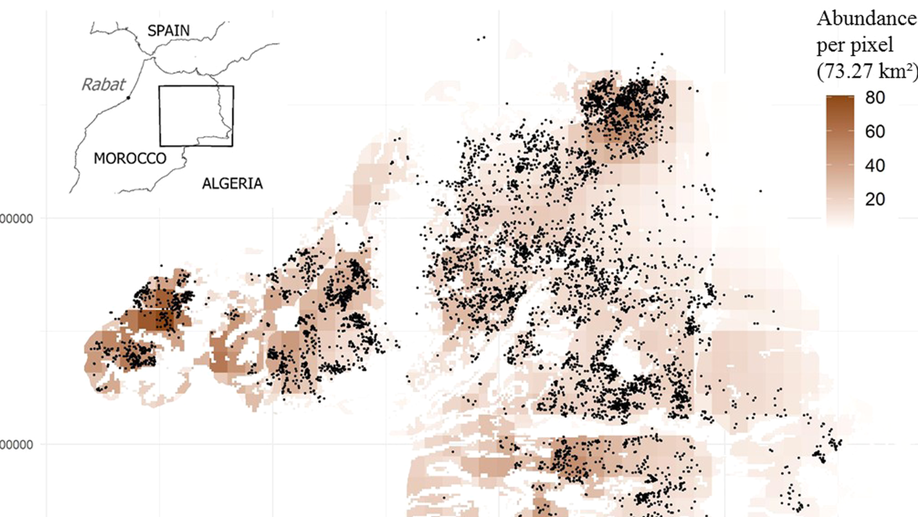
I lead data science teams building production systems at the intersection of geospatial technology and machine learning. My recent work has focused on combining high-resolution Earth observation data with predictive modeling to map soil and crop health at scale, delivering solutions that have secured €2M+ in enterprise contracts and mapped over 200,000 hectares worldwide.
Open Source
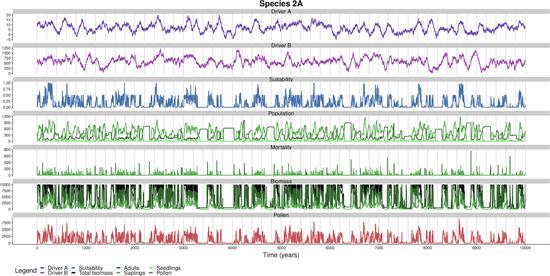
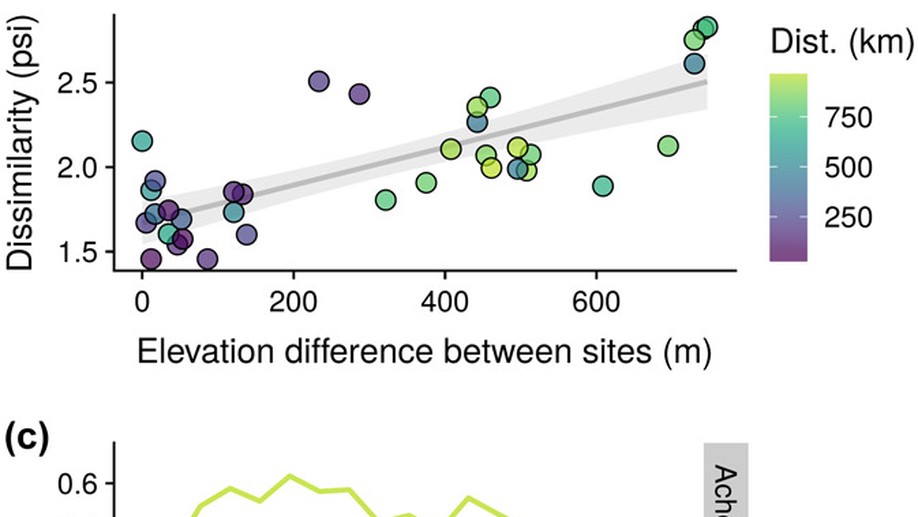
I develop and maintain R packages for spatial and temporal analysis: distantia (dynamic time warping), spatialRF (spatial machine learning), collinear (multicollinearity management), memoria (ecological memory), and virtualPollen (ecological simulation). Together, these packages have been downloaded over 100,000 times.
Research Background
Before transitioning to industry, I built my expertise in world-class research labs across Spain, Denmark, and Norway. I’ve co-authored 49 peer-reviewed papers with 210 collaborators from 22 countries, with several recognized as “Most Downloaded” and “Editor’s Pick” in their respective journals.
Beyond Work
When I’m not working, I enjoy time with my family, tinkering on the piano, paddleboarding, and developing R packages.
Let’s Connect
I’m always open to discussing data science leadership, geospatial technology, and new opportunities. Connect with me on LinkedIn or drop me an email.
Interests
- Geospatial Data Science & Engineering
- Machine Learning & Predictive Modeling
- Earth Observation & Remote Sensing
- Production Systems & Pipeline Automation
- Scientific Software Development
- Team Leadership & Strategic Planning
Education
-
Ph.D. in Computational Ecology, 2010
University of Granada
-
MSc in Geographic Information Systems (UNIGIS), 2009
University of Girona
-
MSc in Management and Environmental Auditing, 2006
University of Cadiz
-
BSc in Biology (Ecology), 2003
University of Granada









{kind=link}
{kind=link}